
文章目录
反二壬烯醛(Trans-2-nonenal 或 E-2-nonenal,以下简称E2N),通常而言被描述为纸板味,是老化啤酒里常常会闻到的味道。当然,你可能并不了解纸板味是什么味道,也不知道为什么老化啤酒会产生它。这是啤酒风味的另一个新专题,让我们认识一下E2N。

总结来说:所谓的纸板味,也就是E2N,它的确是氧化带来的味道,但并不是传统意义上的啤酒储藏导致氧化的味道。
E2N到底是什么味道?
E2N在啤酒领域里通常被认为是“纸板味”(Cardboard Flavor)。但其实它是一些植物的主要气味来源,例如黄瓜。上世纪60到90年代,就有针对黄瓜的气味研究,发现是包含E2N在内的几种醛(Aldehyde),所以很多非啤酒相关的资料会把E2N描述为“黄瓜味”。
除此之外,E2N也是面包味的主要组成部分。这里的E2N主要是美拉德反应的产物。同时研究表明,E2N也是“老人味”的主要味道来源。
E2N的气味阈值过低,容易察觉。根据气味试剂制造商Aroxa提供的数据,E2N的阈值在50-250µg/L。再者,它不算一种令人愉悦的气味,也无法与啤酒相融。因此,才导致大家将其归类为“缺陷”。
尽管讲了黄瓜、纸板、面包甚至老人味,但依旧太抽象,这个时候只能说:大家喝多点就知道了。
——————以下内容较为晦涩——————
E2N为何会出现在啤酒中?
其实关于E2N的研究自千禧年后逐渐减少,因为已经有办法十分有效地控制E2N量,所以学界已经对这个课题不太感兴趣。所以以下结论均为现阶段获得的结果。
- E2N主要产生在制麦和糖化步骤中,之后的酿造步骤里会也会检测到E2N的增加,绝大部分与“壬烯醛潜力”(Nonenal Potential)相关。
- E2N在制麦与糖化过程中增加E2N的机理相差无几。
- E2N的产生机理,分为羟醛缩合反应(Aldol condensation)和氧化,而氧化又分为非酶氧化及酶促氧化。其中酶促氧化反应是E2N来源的大头。
- 酶促反应,其相关的酶是脂氧合酶(Lipoxygenase),在麦芽中是指它的同工酶LOX-1。
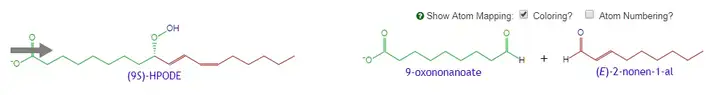
- 反应最核心的步骤是,亚油酸(Linoleic Acid,自然存在或者脂质被脂肪酶降解)在LOX-1的作用下分解为9-HPOD ,再经过“9-HPL Like Activity(9-氢过氧剪切酶样反应)”,降解为E2N。

在看了“每个单字看得明白,但一整句话就不知所谓”的解释后。让我尝试更详细更直白地解释这个原因。
E2N不是一种成品啤酒的“氧化味”
各种科普资料经常含糊地表达纸板味的来源,例如:啤酒在储藏过程中由于啤酒里的溶解氧和未排干净的空气导致啤酒老化,出现纸板味…… 然而实质上,在1999年的一份论文就指出:啤酒储藏中产生的E2N与氧气没有关联。
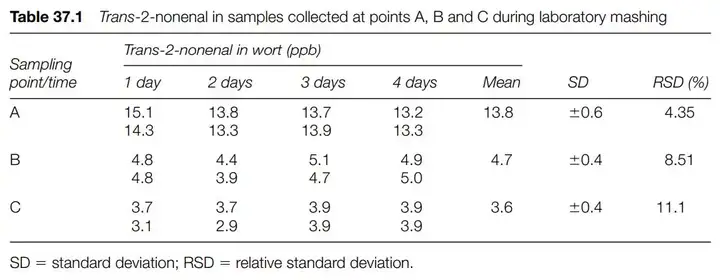
该论文使用一种很暴力的方法,就是往新鲜啤酒通入96ppm的同位素氧气(氧-18),然后观察强制老化(在40摄氏度下存放5日)及自然老化(室温下存放3个月)后,E2N的增长。
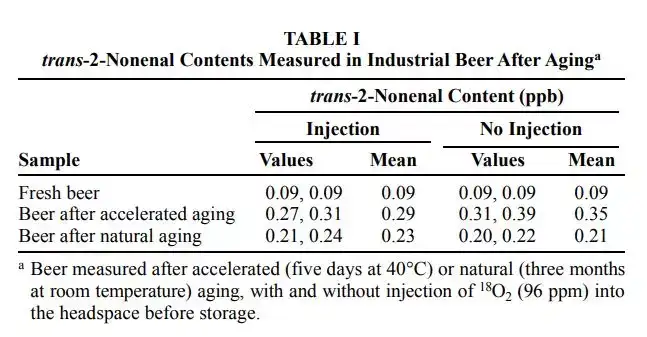
从上面的表格我们可以看出无论注入氧-18与否与E2N的增加并没有关联(Relative)。换言之,在成品啤酒中,氧气的含量与E2N的增加并无关系。那么与什么有关?与壬烯醛潜力相关,这个在下篇文章会介绍。在此之前必须要介绍E2N是如何产生的。
E2N产生机理
E2N在制麦和糖化都有可能会产生。其中的机理大致相同。
让我们追根朔源。在全面研究啤酒的E2N之前,科学家就已经了解到脂质最终会被氧化为各种醛类。最早研究脂质与E2N的关系,来自于各种瓜果蔬菜,黄瓜、甜瓜、香蕉等等。他们了解到这些瓜果的味道是一些醛类物质的表现,尤其是在被切开(或者碾碎)后产生的气味更为明显,这个可能与植物自身防卫机制有关,出于自卫而散发出一些令其他生物抗拒的味道。
在一系列研究后,科学家们得知黄瓜(Cucumis sativus)脂质降解为E2N的过程,中间经历了几个步骤,其中包括酶促氧化、酶促裂合及酶促异构。其步骤如下:
- 脂质被脂肪酶(Lipase,甘油酯水解酶)水解为亚油酸。
- 亚油酸在脂氧合酶的催化下降解为一些氢过氧化合物,主要是13-HOPD及9-HPOD。
- 这些化合物在对应的裂合酶(Lyase)作用下被切割成各种烯醛(Enal),己烯醛,顺-3-壬烯醛(Cis-3-noneal)等。
- 其中顺-3-壬烯醛被烯醛异构酶(Enal Isomerase)异构后得到E2N。
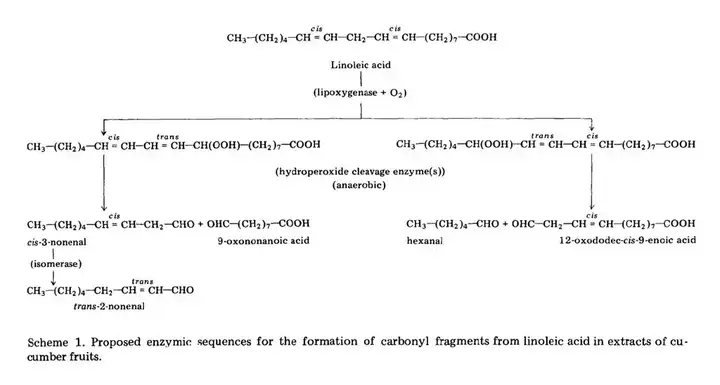
而在大麦的反应中有着与上图几乎相同的途径。
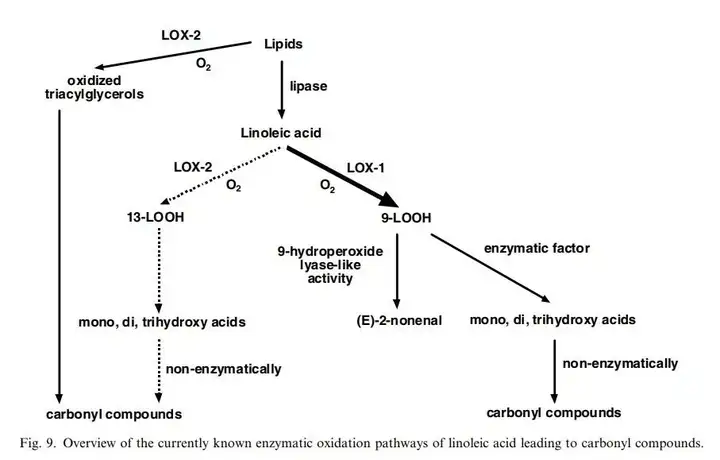
在此解释一下图里的几种化合物,亚油酸是一种双不饱和脂肪酸,大量存在与各种植物中,我们常用的花生油就包含了32%的亚油酸。
9-LOOH以及13-LOOH,即9-HPOD及13-HPOD。9-HPOD((10E,12z)-9-hydroperoxyoctadeca-10,12-dienoic acid,(10E,12Z)-9-氢过氧基十八碳-10,12-二烯酸),是一种亚油酸的代谢物,同样广泛存在于各种生物中。
而所谓的9-HPL like Activity(9-hydroperoxide lyase-like activity),很有可能是9-HPL(9-hydroperoxide lyase,9-氢过氧裂合酶)与烯醛异构酶的共同反应结果。这个将会在下面讲到。
LOX-1
想要从根源性解决问题,就必须从生物的代谢通路入手,下面将解释在通路中遇到的酶和相关的反应。
尽管降解途径看起来与黄瓜的代谢很相似,但不同的生物体的酶尽管功能(也就是基因表达)一样,但基因组成却不尽一样。在大麦里的脂氧合酶是LOX1_HORVU。其中HORVU是指大麦的学名Hordeum vulgare的缩写。部分文献又有HvLox-1的称谓,Hv指的也是学名的缩写。为方便表达,本文还是沿用LOX-1。
LOX-1是一种较为常规的酶,热敏(即高温可以灭活),高活性pH范围为5-8.5。它的主要反应底物(Substrate,指能够催化的化合物)就是亚油酸盐(Linoleate),通过氧促反应代谢为9-HPOD。反应式如下:
Linoleate + O(2) <=> (9S,10E,12Z)-9-hydroperoxy-10,12-octadecadienoate 在路径图里值得注意的是,产物13-HPOD和9-HPOD都是脂氧合酶的同工酶(对应LOX-1,LOX-2)的产物,但是两种同工酶的作用不一样,导致了结果不一样,所以后期的试验中,主要以研究如何减少LOX-1为主。
9-HPL
除了讨论脂氧合酶外,在生成E2N之前还有两个两个重要的步骤,裂合与异构。裂合是9-HPOD降解为醛类的必经步骤。其中为了裂合9-HPOD,有种专门针对它的裂合酶——9-HPL。
9-HPL属于细胞色素P450这个超家族(Super Family,基因超家族,指由同一个基因复制及变异得来的一大组基因),在这个超家族里它称为CYP74C。与CYP74A、CYP74B不一样的是,9-HPL带有HPL基因,所以可以裂合氢过氧化合物。[10]
研究表明,不同植物的9-HPL对于9-HPOD和13-HPOD有着不同的活性,像西瓜就两样都有相同活性,但番石榴的9-HPL就只对9-HPOD有活性。
烯醛异构酶
目前只是假设可能有异构酶的存在。05年,Hisao Kuroda(来自札幌啤酒厂价值创造先锋实验室)检测到在整个9-氢过氧剪切酶样反应,产物从Z3N(3Z-nonenal,即cis-3-nonenal,顺-3-壬烯醛)到E2N的过程。其中,Z代表着顺式(cis),E代表着反式(trans)。
顺式结构的化合物转变为反式结构的化合物,一般顺-反式异构酶(cis-trans isomerae)。而在这个特例里,涉及到的是3Z:2E烯醛异构酶(3Z:2E enal isomerase)。
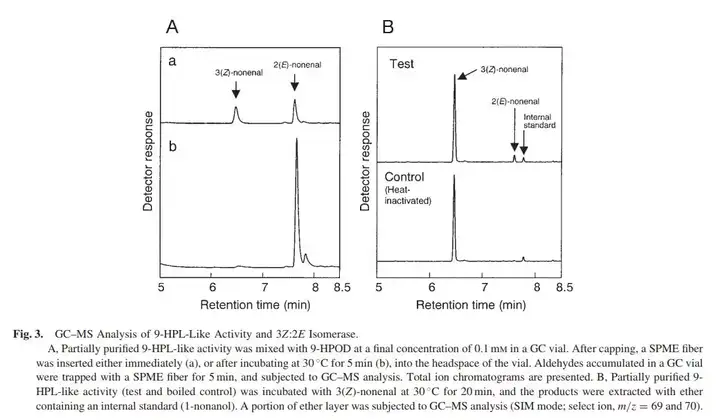
从上图可以看到Z3N在前半段,而E2N位于整个9-氢过氧剪切酶样反应中的后半部分。而成品的反应产物并未检测到Z3N的存在,所以Kuroda就提出类9-HPL活性里包含3Z:2E烯醛异构酶的假设,但试验中无法分离提出异构酶。而他及后来者并没有继续尝试分离。
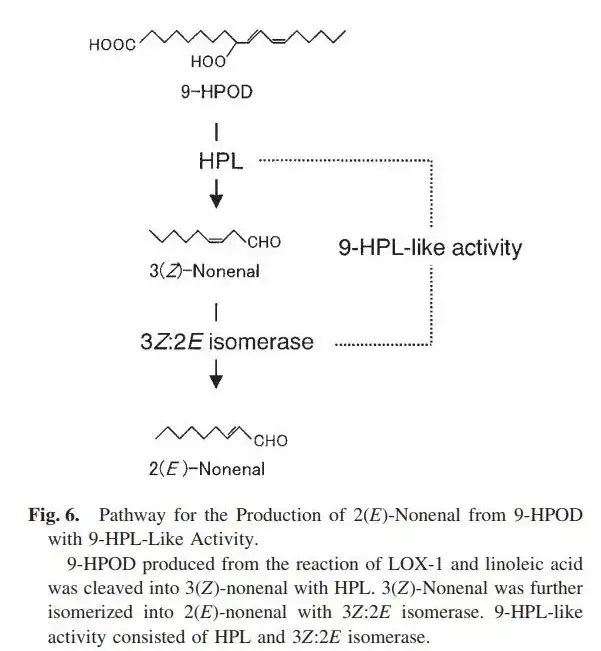
假设这个假说成立,那么这个亚油酸的降解途径就与黄瓜相似了。尽然Kurodo没办法提纯出异构酶,但我们可以用黄瓜的异构酶进行类比。目前所知,黄瓜的3Z:2E烯醛异构酶,pH5.5-9.5时,活性有50%以上。
制麦过程中的E2N产生
制麦(Malting)过程中产生的E2N的机理与糖化一样。只是环境状态不同,产生的量有所不同。
制麦的过程一般分为三个步骤:浸泡(Steeping)、发芽(Germinating)和烘烤(Kilning)。这些一般与家酿爱好者有些距离,但对于大酒厂而言,控制这些变量是必须的。
尽管上面写了很长的关于糖化的E2N反应,但实质上E2N产生最多是在发芽阶段。这些E2N最终会潜伏于麦芽中,从而一直带到成品啤酒里,这就是上面所讲的壬烯醛潜力。我国科研人员通过GC-MS监测从大麦到麦芽的整个发芽过程E2N的变化。可以从下面的表格看到最终发芽时E2N含量很高。

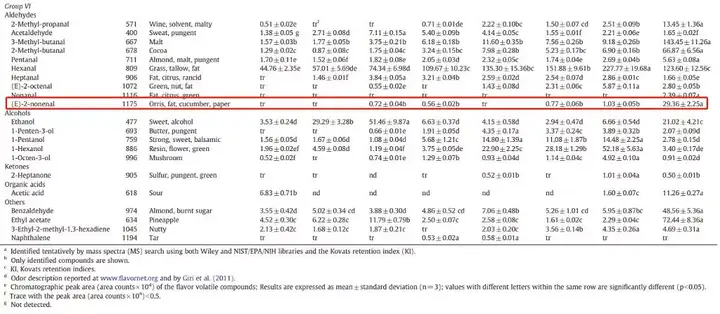
而烘烤的过程,上面就提到LOX-1酶是一种热敏酶,在高温的烘烤环境下酶处于低活性。所以这一个阶段鲜有新的E2N产生(不排除美拉德反应带来新的E2N,但数量不多)。有论文指出在高温加热状态下(尽管论文里是指煮沸麦芽汁的状态),会释放一部分潜伏的E2N,这个我们下一篇细说。
而整个制麦过程的LOX-1酶活性如下图。可以看到LOX-1活性下降了约90%。
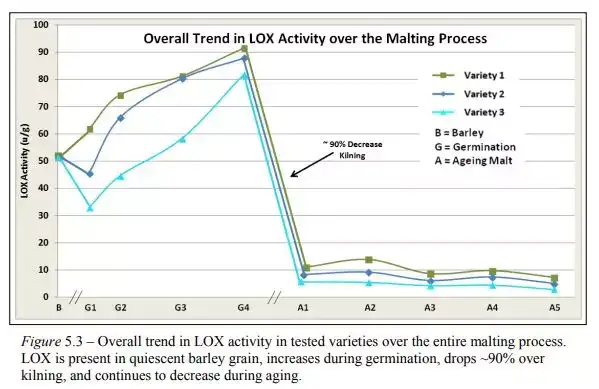
其他可能影响到E2N产生的因素
麦芽品种
不同麦芽品种的LOX-1酶活性不一,甚至9-HPL及顺-反式异构酶活性都不一样。研究人员就尝试收集不同品种的大麦及对应的麦芽来检测E2N含量。不过比较遗憾的是,研究并不是实验者自己去控制变量发芽;而且并未检测壬烯醛潜力值。
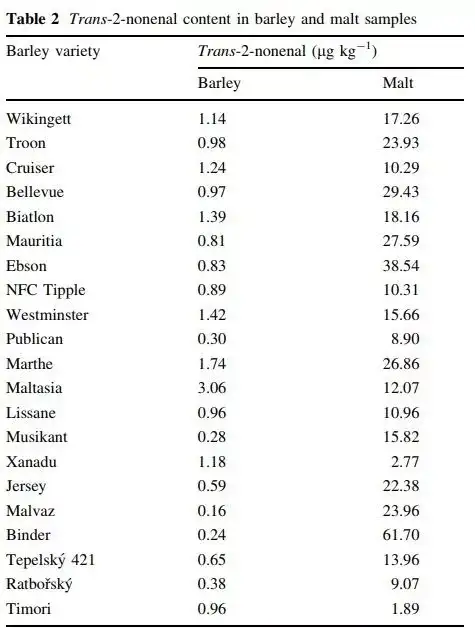
研磨度及麦芽水比
瑞典的研究人员在研究糖化中E2N的变化模型时,将麦芽水比及麦芽研磨度考虑了进去。
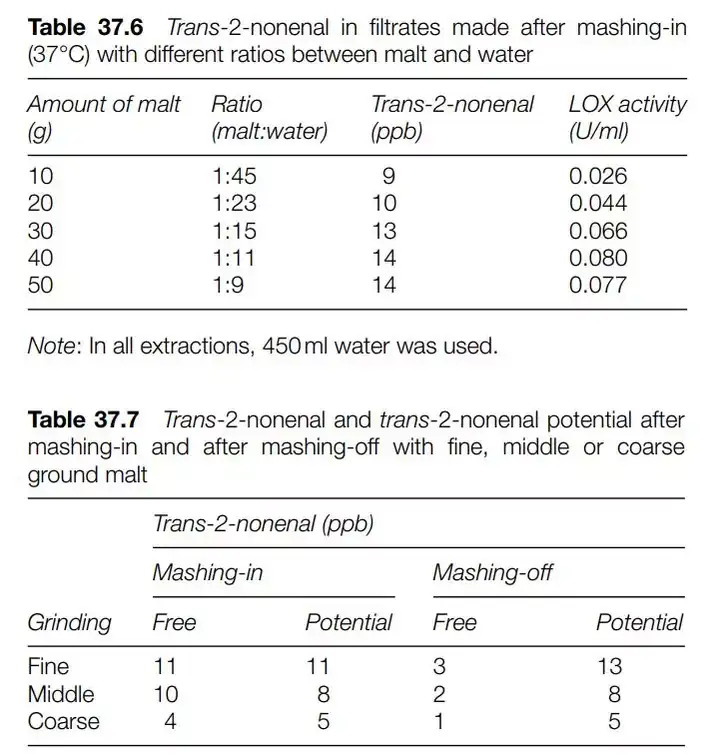
可以观察到麦芽水比到某个阶段后,E2N的含量逐渐稳定,而LOX-1酶活性也趋向稳定。而研磨度越高的麦芽,E2N及壬烯醛潜力更高。
温度与pH值
温度和pH值不仅影响LOX-1的活性,同时影响9-HPL和烯醛异构酶的活性,这里是一个总体的表现。
同样是上面提到的实验人员。他们收集常用的几个糖化温度里的麦芽汁,检测E2N的产生。可以看到温度对E2N的产生影响十分大。
同时他们也做了pH值对E2N产生的影响研究。可以看到pH值越低E2N产生越小。
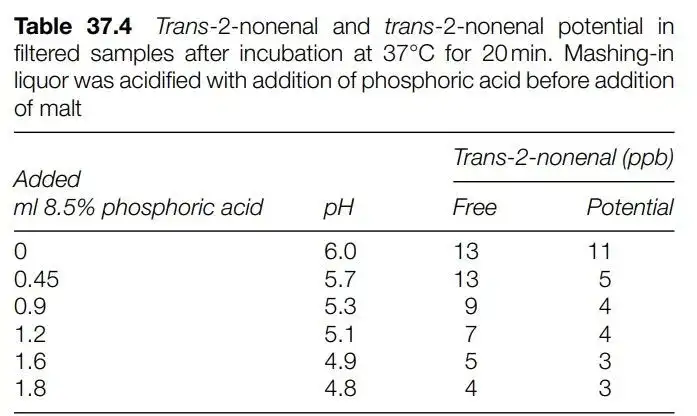
而在实际酿酒操作中:
- 麦芽研磨度一般都在中等范围。
- 麦芽水比为1:1.5到1:5,单位kg/L。
- 糖化温度基本在上述提到的几个温度上下浮动。
- 唯一可以变的是pH值,但pH值牵一发而动全身,况且大部分情况下,糖化的pH都高于5.2。
所以,其实大家并没有太大的空间在常规的酿造过程中控制E2N的数量。所以大家并不需要在意怎么通过提高手法/技艺来避免E2N。
本篇文章主要介绍了E2N的产生机理,和一些影响E2N产生数量的因素。但并未涉及到“解决方案”。下一篇会详细介绍壬烯醛潜力及一些阻止E2N产生的方案。





